Peptidi so razred spojin, ki nastanejo s povezavo več aminokislin s peptidnimi vezmi.V živih organizmih so vseprisotni.Do sedaj so v živih organizmih našli na desettisoče peptidov.Peptidi igrajo pomembno vlogo pri uravnavanju funkcionalnih aktivnosti različnih sistemov, organov, tkiv in celic ter pri življenjskih aktivnostih in se pogosto uporabljajo v funkcionalni analizi, raziskavah protiteles, razvoju zdravil in na drugih področjih.Z razvojem biotehnologije in tehnologije sinteze peptidov je bilo razvitih in uporabljenih v kliniki vse več peptidnih zdravil.
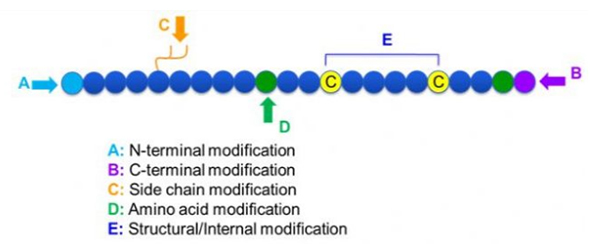
Obstaja veliko različnih modifikacij peptidov, ki jih lahko preprosto razdelimo na post modifikacijo in modifikacijo postopka (z uporabo modifikacije izpeljane aminokisline) in modifikacijo N-terminala, modifikacijo C-terminala, modifikacijo stranske verige, modifikacijo aminokislin, modifikacijo skeleta, itd., odvisno od mesta modifikacije (slika 1).Kot pomembno sredstvo za spremembo strukture glavne verige ali skupin stranskih verig peptidnih verig lahko modifikacija peptidov učinkovito spremeni fizikalne in kemijske lastnosti peptidnih spojin, poveča topnost v vodi, podaljša čas delovanja in vivo, spremeni njihovo biološko porazdelitev, odpravi imunogenost , zmanjšanje toksičnih stranskih učinkov itd. V tem prispevku je predstavljenih več glavnih strategij za spreminjanje peptidov in njihovih značilnosti.
1. Ciklizacija
Ciklični peptidi imajo veliko aplikacij v biomedicini in številni naravni peptidi z biološko aktivnostjo so ciklični peptidi.Ker so ciklični peptidi bolj togi kot linearni peptidi, so izjemno odporni na prebavni sistem, lahko preživijo v prebavnem traktu in kažejo večjo afiniteto za ciljne receptorje.Ciklizacija je najbolj neposreden način za sintezo cikličnih peptidov, zlasti za peptide z velikim strukturnim skeletom.Glede na način ciklizacije ga lahko razdelimo na tip stranske verige stranske verige, tip terminala - stranske verige, tip terminala - terminala (tip od konca do konca).
(1) od stranske verige do stranske verige
Najpogostejši tip ciklizacije stranske verige v stransko verigo je disulfidni most med cisteinskimi ostanki.To ciklizacijo uvede par cisteinskih ostankov, ki so deprotektirani in nato oksidirani, da tvorijo disulfidne vezi.Policiklično sintezo lahko dosežemo s selektivnim odstranjevanjem sulfhidrilnih zaščitnih skupin.Ciklizacijo lahko izvedemo v postdisociacijskem topilu ali na preddisociacijski smoli.Ciklizacija na smolah je lahko manj učinkovita kot ciklizacija s topilom, ker peptidi na smolah ne tvorijo zlahka ciklificiranih konformacij.Druga vrsta ciklizacije stranske verige je tvorba amidne strukture med ostankom asparaginske ali glutaminske kisline in osnovno aminokislino, kar zahteva, da je zaščitno skupino stranske verige mogoče selektivno odstraniti iz polipeptida bodisi na smoli ali po disociaciji.Tretja vrsta stranske verige - ciklizacija stranske verige je tvorba difenil etrov s tirozinom ali p-hidroksifenilglicinom.To vrsto ciklizacije v naravnih izdelkih najdemo le v mikrobnih izdelkih, izdelki ciklizacije pa imajo pogosto potencialno medicinsko vrednost.Priprava teh spojin zahteva edinstvene reakcijske pogoje, zato se pogosto ne uporabljajo pri sintezi običajnih peptidov.
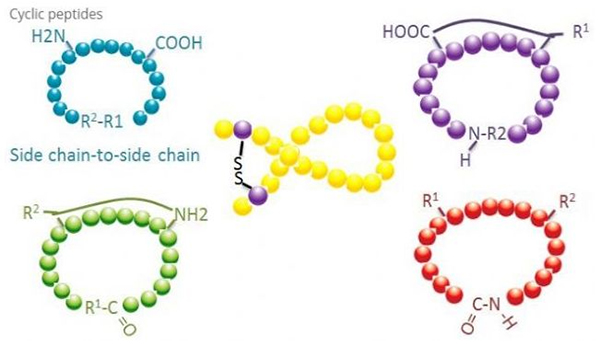
(2) od terminala do stranske verige
Ciklizacija končne stranske verige običajno vključuje C-terminal z amino skupino lizinske ali ornitinske stranske verige ali N-terminal s stransko verigo asparaginske ali glutaminske kisline.Druga polipeptidna ciklizacija poteka z tvorbo etrskih vezi med terminalom C in stranskimi verigami serina ali treonina.
(3) Končni tip ali tip od glave do repa
Verižne polipeptide je mogoče ciklirati v topilu ali fiksirati na smolo s cikliranjem stranske verige.Pri centralizaciji topila je treba uporabiti nizke koncentracije peptidov, da se izognemo oligomerizaciji peptidov.Izkoristek sintetičnega obročnega polipeptida od glave do repa je odvisen od zaporedja verižnega polipeptida.Zato je treba pred pripravo cikličnih peptidov v velikem obsegu najprej ustvariti knjižnico možnih verižnih vodilnih peptidov, čemur sledi ciklizacija, da se najde zaporedje z najboljšimi rezultati.
2. N-metilacija
N-metilacija se prvotno pojavi v naravnih peptidih in se vnese v sintezo peptidov, da se prepreči tvorba vodikovih vezi, zaradi česar so peptidi bolj odporni na biorazgradnjo in očistek.Najpomembnejša metoda je sinteza peptidov z uporabo N-metiliranih derivatov aminokislin.Poleg tega se lahko uporabi tudi Mitsunobujeva reakcija intermediatov N-(2-nitrobenzen sulfonil klorid) polipeptid-smola z metanolom.Ta metoda je bila uporabljena za pripravo cikličnih peptidnih knjižnic, ki vsebujejo N-metilirane aminokisline.
3. Fosforilacija
Fosforilacija je ena najpogostejših posttranslacijskih sprememb v naravi.V človeških celicah je več kot 30 % beljakovin fosforiliranih.Fosforilacija, zlasti reverzibilna fosforilacija, ima pomembno vlogo pri nadzoru številnih celičnih procesov, kot so prenos signala, izražanje genov, regulacija celičnega cikla in citoskeleta ter apoptoza.
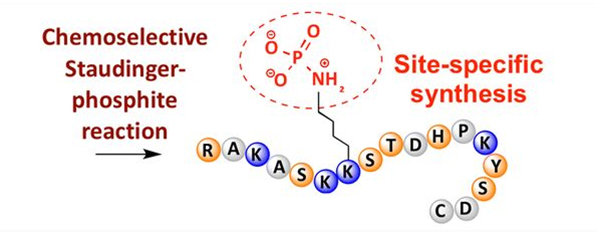
Fosforilacijo lahko opazimo pri različnih aminokislinskih ostankih, vendar so najpogostejše tarče fosforilacije ostanki serina, treonina in tirozina.Derivati fosfotirozina, fosfotreonina in fosfoserina se lahko vnesejo v peptide med sintezo ali pa nastanejo po sintezi peptida.Selektivno fosforilacijo lahko dosežemo z uporabo ostankov serina, treonina in tirozina, ki selektivno odstranijo zaščitne skupine.Nekateri fosforilacijski reagenti lahko s post modifikacijo v polipeptid vnesejo tudi skupine fosforne kisline.V zadnjih letih je bila fosforilacija lizina, specifična za mesto, dosežena s kemično selektivno reakcijo Staudinger-fosfit (slika 3).
4. Miristoilacija in palmitoilacija
Acilacija N-terminala z maščobnimi kislinami omogoča, da se peptidi ali proteini vežejo na celične membrane.Miridamoilirano zaporedje na N-terminalu omogoča ciljno vezavo protein kinaz družine Src in proteinov Gaq reverzne transkriptaze na celične membrane.Miristično kislino smo povezali z N-terminalom smolnega polipeptida z uporabo standardnih reakcij pripajanja in nastali lipopeptid je bilo mogoče disociirati pod standardnimi pogoji in očistiti z RP-HPLC.
5. Glikozilacija
Glikopeptidi, kot sta vankomicin in teikolanin, so pomembni antibiotiki za zdravljenje bakterijskih okužb, odpornih na zdravila, drugi glikopeptidi pa se pogosto uporabljajo za stimulacijo imunskega sistema.Poleg tega je veliko mikrobnih antigenov glikoziliranih, zato je zelo pomembno proučevanje glikopeptidov za izboljšanje terapevtskega učinka okužbe.Po drugi strani pa je bilo ugotovljeno, da proteini na celični membrani tumorskih celic kažejo nenormalno glikozilacijo, zaradi česar imajo glikopeptidi pomembno vlogo pri raziskavah raka in tumorske imunske obrambe.Glikopeptide pripravimo po metodi Fmoc/t-Bu.Glikozilirane ostanke, kot sta treonin in serin, pogosto vnašajo v polipeptide fMOC-ji, aktivirani s pentafluorofenolnim estrom, da zaščitijo glikozilirane aminokisline.
6. Izopren
Do izopentadienilacije pride na cisteinskih ostankih v stranski verigi blizu C-terminala.Proteinski izopren lahko izboljša afiniteto celične membrane in tvori interakcijo beljakovine in beljakovine.Izopentadienirani proteini vključujejo tirozin fosfatazo, majhno GTazo, molekule kohaperona, jedrno lamino in centromerne vezavne proteine.Izoprenske polipeptide lahko pripravimo z uporabo izoprena na smolah ali z uvedbo derivatov cisteina.
7. Modifikacija polietilen glikola (PEG).
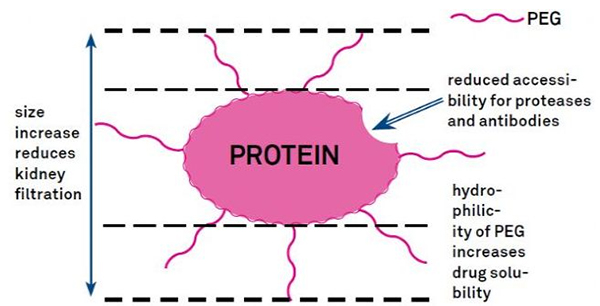
Modifikacijo PEG lahko uporabimo za izboljšanje hidrolitske stabilnosti beljakovin, biološke porazdelitve in topnosti peptidov.Uvedba verig PEG v peptide lahko izboljša njihove farmakološke lastnosti in tudi zavre hidrolizo peptidov s proteolitičnimi encimi.Peptidi PEG prehajajo skozi presek glomerularnih kapilar lažje kot običajni peptidi, kar močno zmanjša ledvični očistek.Zaradi podaljšane aktivne razpolovne dobe peptidov PEG in vivo je mogoče normalno raven zdravljenja vzdrževati z nižjimi odmerki in manj pogostimi peptidnimi zdravili.Vendar pa ima modifikacija PEG tudi negativne učinke.Velike količine PEG preprečujejo, da bi encim razgradil peptid, poleg tega pa zmanjšajo vezavo peptida na tarčni receptor.Toda nizka afiniteta peptidov PEG je običajno izravnana z njihovim daljšim farmakokinetičnim razpolovnim časom in ker so prisotni v telesu dlje, je verjetnost, da se peptidi PEG absorbirajo v ciljna tkiva, večja.Zato je treba specifikacije polimera PEG optimizirati za optimalne rezultate.Po drugi strani pa se PEG peptidi kopičijo v jetrih zaradi zmanjšanega ledvičnega očistka, kar povzroči makromolekularni sindrom.Zato je treba modifikacije PEG načrtovati bolj natančno, ko se peptidi uporabljajo za testiranje zdravil.
Pogoste modifikacijske skupine modifikatorjev PEG lahko v grobem povzamemo kot sledi: amino (-amin) -NH2, aminometil-Ch2-NH2, hidroksi-OH, karboksi-Cooh, sulfhidril (-tiol) -SH, maleimid -MAL, sukcinimid karbonat - SC, sukcinimid acetat -SCM, sukcinimid propionat -SPA, n-hidroksisukcinimid -NHS, akrilat-ch2ch2cooh, aldehid -CHO (kot je propional-ald, butyrALD), akrilna baza (-akrilat-acrl), azido-azid, biotinil - Biotin, fluorescein, glutaril -GA, akrilat hidrazid, alkin-alkin, p-toluensulfonat -OTs, sukcinimid sukcinat -SS itd. Derivati PEG s karboksilnimi kislinami se lahko sklopijo na n-terminalne amine ali stranske verige lizina.PEG, aktiviran z aminokislinami, lahko povežemo s stranskimi verigami asparaginske kisline ali glutaminske kisline.Nepravilno aktiviran PEG je mogoče konjugirati v merkaptan s popolnoma nezaščitenimi stranskimi verigami cisteina [11].Modifikatorji PEG so običajno razvrščeni kot sledi (opomba: mPEG je metoksi-PEG, CH3O-(CH2CH2O)n-CH2CH2-OH):
(1) ravnoverižni modifikator PEG
mPEG-SC, mPEG-SCM, mPEG-SPA, mPEG-OTs, mPEG-SH, mPEG-ALD, mPEG-butyrALD, mPEG-SS
(2) bifunkcionalni modifikator PEG
HCOO-PEG-COOH, NH2-PEG-NH2, OH-PEG-COOH, OH-PEG-NH2, HCl·NH2-PEG-COOH, MAL-PEG-NHS
(3) modifikator razvejanja PEG
(mPEG)2-NHS, (mPEG)2-ALD, (mPEG)2-NH2, (mPEG)2-MAL
8. Biotinizacija
Biotin se lahko močno veže z avidinom ali streptavidinom, vezavna moč pa je celo blizu kovalentni vezi.Peptidi, označeni z biotinom, se pogosto uporabljajo v imunskih testih, histocitokemiji in pretočni citometriji na osnovi fluorescence.Označena protitelesa proti antibiotikom se lahko uporabljajo tudi za vezavo biotiniliranih peptidov.Biotinske oznake so pogosto pritrjene na stransko verigo lizina ali N terminal.6-aminokaprojska kislina se pogosto uporablja kot vez med peptidi in biotinom.Vez je prožna pri vezavi na substrat in se bolje veže v prisotnosti sterične ovire.
9. Fluorescentno označevanje
Fluorescentno označevanje se lahko uporablja za sledenje polipeptidom v živih celicah ter za preučevanje encimov in mehanizmov delovanja.Triptofan (Trp) je fluorescenten, zato se lahko uporablja za intrinzično označevanje.Emisijski spekter triptofana je odvisen od perifernega okolja in pada z zmanjševanjem polarnosti topila, lastnost, ki je uporabna za odkrivanje peptidne strukture in vezave na receptor.Fluorescenco triptofana je mogoče ugasniti s protonirano asparaginsko kislino in glutaminsko kislino, kar lahko omeji njegovo uporabo.Dansil kloridna skupina (Dansyl) je močno fluorescentna, ko je vezana na amino skupino in se pogosto uporablja kot fluorescentna oznaka za aminokisline ali beljakovine.
Fluorescenčna resonanca Pretvorba energije (FRET) je uporabna za študije encimov.Ko se uporabi FRET, substratni polipeptid običajno vsebuje skupino za označevanje fluorescence in skupino za dušenje fluorescence.Označene fluorescentne skupine duši dušilec z nefotonskim prenosom energije.Ko je peptid disociiran od zadevnega encima, skupina za označevanje oddaja fluorescenco.
10. Polipeptidi kletke
Peptidi v kletki imajo optično odstranljive zaščitne skupine, ki ščitijo peptid pred vezavo na receptor.Ko je izpostavljen UV-sevanju, se peptid aktivira in obnovi svojo afiniteto do receptorja.Ker je to optično aktivacijo mogoče nadzorovati glede na čas, amplitudo ali lokacijo, se lahko peptidi v kletki uporabljajo za preučevanje reakcij, ki se pojavljajo v celicah.Najpogosteje uporabljene zaščitne skupine za polipeptide v kletki so 2-nitrobenzilne skupine in njihovi derivati, ki jih je mogoče v peptidno sintezo uvesti preko zaščitnih derivatov aminokislin.Razviti derivati aminokislin so lizin, cistein, serin in tirozin.Derivati aspartata in glutamata pa se običajno ne uporabljajo zaradi njihove dovzetnosti za ciklizacijo med sintezo in disociacijo peptidov.
11. Poliantigenski peptid (MAP)
Kratki peptidi običajno niso imuni in jih je treba povezati z nosilnimi proteini, da proizvedejo protitelesa.Poliantigenski peptid (MAP) je sestavljen iz več identičnih peptidov, povezanih z lizinskimi jedri, ki lahko specifično izražajo imunogene z visoko močjo in se lahko uporabijo za pripravo sklopov peptid-nosilni protein.Polipeptide MAP lahko sintetiziramo s sintezo v trdni fazi na smoli MAP.Vendar nepopolno spajanje povzroči manjkajoče ali okrnjene peptidne verige na nekaterih vejah in tako ne kaže lastnosti prvotnega polipeptida MAP.Kot alternativo lahko peptide pripravimo in očistimo ločeno ter jih nato povežemo z MAP.Peptidno zaporedje, vezano na peptidno jedro, je dobro definirano in ga je enostavno karakterizirati z masno spektrometrijo.
Zaključek
Peptidna modifikacija je pomembno sredstvo za oblikovanje peptidov.Kemično modificirani peptidi ne morejo samo vzdrževati visoke biološke aktivnosti, temveč se tudi učinkovito izogniti pomanjkljivostim imunogenosti in toksičnosti.Hkrati lahko kemična modifikacija peptidom podeli nekatere nove odlične lastnosti.V zadnjih letih se je metoda aktivacije CH za postmodifikacijo polipeptidov hitro razvijala in dosegli so številne pomembne rezultate.
Čas objave: 20. marec 2023